HI-Virus










Trainier deine Lernmuskeln!
Mit Flash Cards, Quiz und mehr
LoslegenAbkürzung: HIV (Human Immunodeficiency Virus)
Synonyme: LAV (Lymphadenopathie-assoziiertes Virus), HTLV III (Humanes T-Zell-lymphotropes Virus III), ARV (AIDS-assoziiertes Retrovirus), AIDS-Virus, häufig fälschlicherweise auch HIV-Virus
Englisch: HIV
Definition
Das HI-Virus, kurz HIV, ist ein RNA-Virus, das zur Gruppe der Lentivirinae innerhalb der Familie der Retroviridae zählt. Es verursacht die HIV-Infektion, die ohne Behandlung im Spätstadium meist zu AIDS führt.
Einteilung
Bisher sind zwei Typen von HI-Viren bekannt, das HIV-1 (inklusive Subtyp 0) mit den Subtypen M (Major), N (New) und O (Outlier) sowie das HIV-2, das im Wesentlichen in Westafrika endemisch ist und eine geringere Pathogenität aufweist. HIV-1 tritt sowohl global als auch in Deutschland deutlich häufiger auf als HIV-2. Deshalb liegt der Schwerpunkt der pharmakologischen Forschung auf dieser Virusvariante. Unter HIV-1 ist der M-Subtyp weltweit der häufigste.
Das HI-Virus gehört neben SARS-CoV-2 zu den am besten untersuchten und erforschten Viren überhaupt.
Geschichte
Über die Geschichte des HI-Virus berichten die meisten Quellen uneinheitlich. Die am weitesten akzeptierte Lehrmeinung beinhaltet die Übertragung des Virus von Primaten auf den Menschen im Laufe der ersten Hälfte des 20. Jahrhunderts. Grund für diese Annahme ist die Ähnlichkeit zu in diesen Tieren vorkommenden Viren, vor allem dem SI-Virus (Simian Immunodeficiency Virus), das in Primaten Symptome auslöst, die denen von mit HIV infizierten Menschen ähneln.
Allgemein wird angenommen, dass die ersten Fälle von HIV-Infektionen in Afrika auftraten; von dort aus lässt sich die Erkrankung nach Haiti und später in die USA verfolgen, wo erste HIV-Fälle 1981 beschrieben wurden. Von ersten Infektionen in Europa wurde 1982 berichtet. Im Nachhinein untersuchte Blutproben wiesen HIV-Fragmente in Blutproben nach, die 1959 im Kongo entnommen wurden.
Die erste Isolierung des HI-Virus gelang Luc Montagnier 1983 aus einem Patienten mit Symptomen einer Lymphadenopathie; er nannte das Virus LAV (lymphadenopathia-associated virus). Dasselbe Virus wurde Monate später von Robert Gallo erneut isoliert, der es aufgrund seiner Ähnlichkeit zu den bereits bekannten lymphotropen Retroviren als HTLV-3 ("human T-lymphotropic virus") bezeichnete. Zwischen beiden Forschern entbrannte ein Streit um die Erstbeschreibung des Erregers, der erst durch eine gemeinsame Konferenz in Paris beigelegt werden konnte.
Erste Testmöglichkeiten gegen HIV kamen 1985 in Umlauf; sie basierten auf dem Nachweis von virusspezifischen Antikörpern im Blut. Dadurch wurde die Testung von Blutprodukten auf Virusbefall möglich.
Die Bindung des Virus an den CD4-Rezeptor von T-Zellen und Makrophagen wurde noch im selben Jahr nachgewiesen. 1986 entdeckte man das Typ-II-HIV und stellte mit dem AZT das erste lebensverlängernde Medikament vor.
Seit 1996 ist die Kombinationstherapie mit mehreren Virostatika Standard in der Behandlung von AIDS.
Genetik
Das Genom des HI-Virus enthält nur neun Gene, so dass das Virus bei Replikation und Transkription seiner Proteine weitgehend auf zelluläre Enzyme und Substrate angewiesen ist.
Aufbau der viralen Nukleinsäure
Das Virusgenom besteht aus zwei Kopien einer Plusstrang-RNA-Kette, die im Core mit Proteinen assoziiert vorliegt, jedoch bei der Transkription nicht direkt als mRNA fungieren kann.
Am 5'- und 3'-Ende der RNA befinden sich sogenannte LTR-Regionen (long terminal repeats), die bei der Bildung viraler Genprodukte als starke Promotoren wirken; dazwischen befinden sich die Gene des HI-Virus, die für alle strukturellen und enzymatisch wirksamen Proteine kodieren.
Genom
Die RNA des HI-Virus weist neun Gene auf; davon kodieren lediglich drei Gene (gag, pol, env) für weitergegebene Proteine, die anderen sechs Gene (Rev, Nef, Tat, Vif, Vpr, Vpu) kodieren für Regulationsfaktoren genetischer Prozesse.
- gag (group specific antigen): Das gag-Gen kodiert für die Proteine des Kapsids (p7, p24 und p17).
- pol (polymerase): Die Information für alle enzymatischen Proteine des Virus, für reverse Transkriptase, Protease und Integrase, befindet sich im Bereich des pol-Gens.
- env (envelope): Das env-Gen beinhaltet die Erbinformation für die beiden Glykoproteine der Virushülle, gp41 und gp120.
- rev (regulator of virion): rev stimuliert die Expression viraler Strukturproteine.
- tat (transactivation of transcription): Das tat-Gen stimuliert die LTR-vermittelte Produktion von viraler RNA.
- vif (virion infectivity factor): vif unterstützt die Bildung infektiöser HI-Viren
- vpr (viral protein r): vpr induziert einen G2/M-Zellzyklusarrest der Wirtszelle und erleichtert den nukleären Import des Präintegrationskomplexes
- vpu (viral protein u) vpu ist essentiell für die Virusreifung und Freisetzung
- nef (negative factor) nef fördert die Virusreplikation und Immunevasion durch Downregulation von CD4- und MHC-I-Molekülen auf der Oberfläche infizierter Zellen
Mutationen
Das HI-Virus besitzt nur einen Teil der Proteinausstattung, die für den Ablauf des viralen Vermehrungszyklus benötigt wird; es ist daher auf den Replikations- und Transkriptionsapparat der Wirtszelle angewiesen.
Nach Infektion einer Zelle wird das RNA-Genom des Virus in DNA transkribiert und in das Genom der Wirtszelle integriert.
Sowohl die reverse Transkriptase als auch die menschlichen RNA-Polymerasen bauen statistisch gesehen etwa jede 10.000te Base fehlerhaft ein. Da beide Enzyme keine Möglichkeit von Proofreading oder Fehlpaarungsreparatur haben, verursachen sie damit mit einer Wahrscheinlichkeit von 10-4 fehlerhafte Informationen, so dass jede neu synthetisierte Nukleinsäure statistisch zwischen 3 und 5 Fehler enthält. Auf diese Weise entstehen mit der Zeit diverse Mutanten des HI-Virus, von denen einige vermehrungsfähig sind und andere Zellen infizieren. Sie weisen aber Unterschiede in der Aminosäuresequenz ihrer Proteine auf: Im Laufe der HIV-Erkrankung verändert sich damit die Struktur der Viren kontinuierlich und entzieht sich unter anderem so einer effektiven Immunabwehr.
Genom-Mutationen können zu insgesamt aggressiveren Virus-Varianten führen. Anfang des Jahres 2005 wurde in New York bei einem HIV-Patienten eine neue Variante des HI-Virus gefunden, die einen deutlich schnelleren Erkrankungsverlauf verursacht und eine multiple Resistenz gegen HIV-Therapeutika besitzt ("Super-HIV"). Diese Virus-Mutation scheint die Zytokinrezeptoren CCR5 und CXCR4 (X4) gleichzeitig für die Virusbindung nutzen zu können.
Proteine
Neben den oben erläuterten regulatorischen Proteinen kodiert das Virusgenom vor allem für Strukturproteine und drei Enzyme, die nach der Freisetzung des Virus Bestandteil des Partikels sind.
Die strukturellen Proteine tragen Abkürzungen (p für Protein, gp für Glykoprotein) und Nummern, die das Molekulargewicht in Kilodalton (kDa) angeben:
- Strukturproteine (kodiert in den Genen gag und env)
- p17: p17 ist ein unterhalb der Virushülle befindliches Matrixprotein.
- p24: p24 bildet das Kapsid.
- p7: p7 bindet an die RNA.
- gp41: Das aus dem Vorläuferprotein gp160 zusammen mit gp120 gebildete Glykoprotein durchzieht transmembranär die Virushülle.
- gp120: Glykoprotein 120 ist mit gp41 assoziiert, mit dem zusammen es transkribiert wird und befindet sich auf der Außenseite der Virushülle.
- Enzyme (kodiert im pol-Gen)
- p66 (reverse Transkriptase): Die reverse Transkriptase ist in der Lage, entgegen dem zentralen Dogma der Zellbiologie RNA in DNA zu transkribieren. Dabei wird der virale RNA-Strang in komplementäre DNA (cDNA) umgeschrieben. Durch die RNA-DNA-Polymerase-Aktivität der reversen Transkriptase wird aus der cDNA die Doppelstrang-DNA gebildet. Durch die RNase H wird der RNA-Strang abgebaut.
- p11 (Protease): Die Protease katalysiert die hydrolytische Spaltung der viralen Proteine, die zu mehreren in funktionellen Einheiten translatiert werden. Von der Polypeptidkette, die vom pol-Gen kodiert wird, spaltet die Protease erst sich selbst ab und trennt danach die reverse Transkriptase von der Integrase.
- p32 (Integrase): Die Integrase ist in der Lage, den von der reversen Transkriptase synthetisierten DNA-Doppelstrang in das Genom der Wirtszelle einzubauen.
Morphologie
Das HI-Virus hat einen Durchmesser von etwa 100 nm und erscheint auf elektronenmikroskopischen Bildern rundlich.
Nukleokapsid (Core)
Im Zentrum des Virus befinden sich zwei Kopien des Virus-Genoms in Form einzelsträngiger RNA-Moleküle. An die RNA sind mehrere Proteine angelagert, hierzu zählt primär das Nukleokapsid-Protein p7. Ebenfalls mit der Nukleinsäure assoziiert sind die reverse Transkriptase, die Integrase sowie Rev und Tat.
Das Genom des HI-Virus ist von einem kegelförmigen Kapsid eingeschlossen. Dieses besteht aus etwa 200 hexameren und 12 pentameren Kapsomeren des Kapsidproteins p24.
Hülle
Bei der Freisetzung des Virus aus der Wirtszelle umgibt sich das Kapsid typischerweise mit einer Virushülle aus Phospholipiden der Wirtszellmembran. An diese Membranlipide ist von innen eine Schicht aus viralen Matrixproteinen gebunden, die hauptsächlich aus p17-Trimeren besteht. Zwischen Hülle und Kapsid befinden sich weitere Proteine, z.B. die Protease (p11), Vpr, Vif und Nef.
In der Lipiddoppelschicht sind neben zellulären Membranproteine (z.B. humanes HLA) zwei virale Glykoproteine verankert: das Transmembranprotein gp41 und das extrazelluläre Protein gp120, welches für das Eindringen in die Wirtszellen von Bedeutung ist.
Übertragung
Das HIV ist sehr empfindlich gegenüber Luftexposition und Desinfektionsmitteln, so dass eine Übertragung durch Tröpfchen- oder Schmierinfektion nahezu ausgeschlossen ist.
Vor allem ein direkter Kontakt mit Körperflüssigkeiten, vorwiegend Blut und Genitalsekreten, führt (allerdings statistisch gesehen nur in seltenen Fällen) zu einer Aufnahme des Virus und damit zu einer HIV-Infektion.
Typische Übertragungswege beinhalten den Geschlechtsverkehr, Gebrauch von infizierten Kanülen (Nadelstichverletzung) und medizinischen Geräten, intravenöse Übertragung von Blutprodukten und die perinatale Weitergabe.
Epidemiologie
siehe: AIDS - Epidemiologie
Infektionsablauf
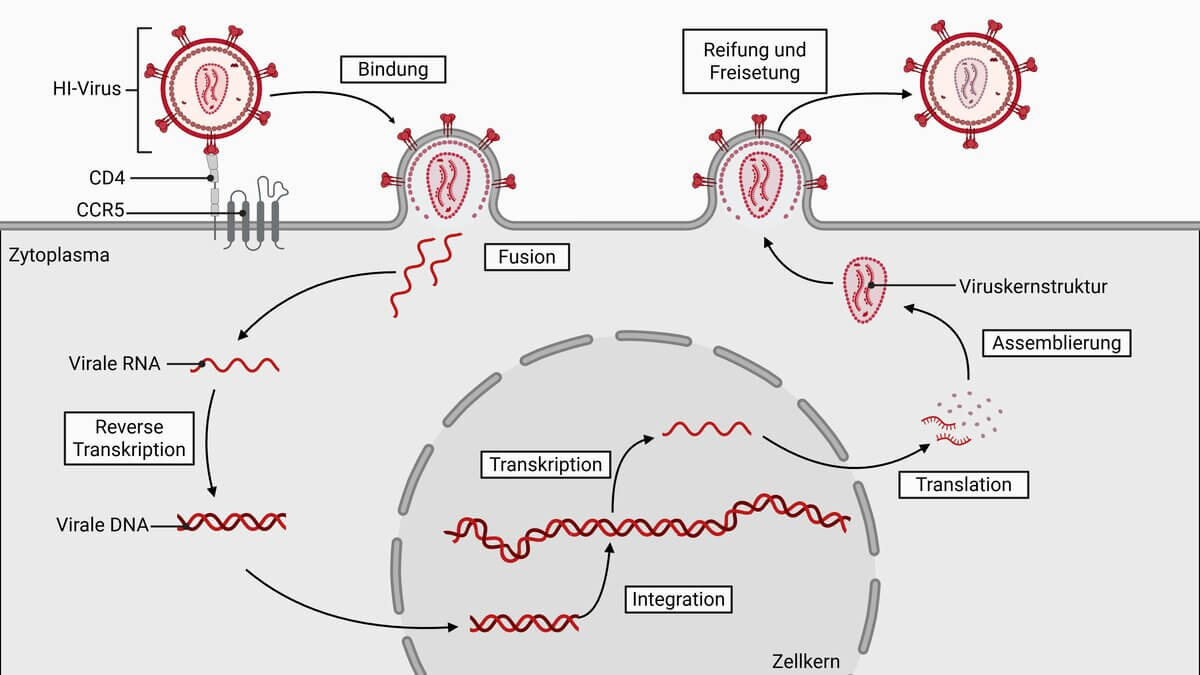
Nach heutigem Wissensstand (2024) gliedert sich der virale Zyklus in sieben Phasen, die das Virus von Befall der Wirtszelle bis zu seiner erneuten Freisetzung durchlaufen muss.
Virusadsorption
Nach dem Eindringen des Virus in die Blutbahn des Menschen hat das Virus die Möglichkeit, über sein Oberflächenprotein gp120 an den CD4-Rezeptor verschiedener Blutzellen zu binden, vor allem CD4+-T-Zellen und Makrophagen. Prinzipiell sind jedoch alle Zellen, die den CD4-Rezeptor exprimieren, potentielle Zielzellen des Virus; dies trifft damit auch auf bestimmte Mikroglia-Zellen des ZNS und Zellen des Gastrointestinaltraktes zu.
Virusfusion
Nach der Bindung des HI-Virus an seine Wirtszelle bildet sich innerhalb der Zellmembran eine Einbuchtung aus. Durch Vermittlung weiterer Rezeptoren, vor allem der Zytokinrezeptoren CCR5 und CXCR4, kann das Virus fester binden und seine Membran mit der Lipiddoppelschicht der Wirtszelle fusionieren. Dabei wird das Kapsid in das Zytosol freigesetzt.
Freisetzung der Virus-RNA
In einem als Uncoating bezeichneten Vorgang, setzt das Kapsid innerhalb des Zytoplasmas der Wirtszelle das Virusgenom frei. Die dabei relevanten Mechanismen sind noch weitgehend unverstanden.
- Anmerkung: Es ist bekannt, dass sich das HI-Virus in den meisten Primaten nicht replizieren kann. Sie haben durch die Expression des Proteins TRIM 5α einen natürlichen Schutz vor HIV, nicht aber vor SIV, einem HIV-ähnlichen Virus, das Affen befällt.
Reverse Transkription
Zusammen mit der Virus-RNA werden auch die im Viruskapsid enthaltenen Proteine freigesetzt. Die in die Zelle eingeführte reverse Transkriptase beginnt unmittelbar mit der Synthese des komplementären DNA-Stranges und danach mit der Bildung von DNA-Doppelsträngen, die sich zu Ringen schließen. Als Primer benutzt sie dabei menschliche tRNA-Moleküle, die im Zytoplasma vorhanden sind oder als Bestandteile des Kapsids aus lysierten Zellen mitgenommen wurden.
- Anmerkung: Auch wenn das Erbgut des HI-Virus aus Plusstrang-RNA besteht, kann es dennoch nicht als Matrize für die Proteinbiosynthese dienen (mRNA), sondern muss erst in DNA transkribiert und ins Genom der Wirtszelle integriert werden.
DNA-Integration
Die ringförmigen DNA-Doppelstränge werden durch das Enzym Integrase katalysiert und vermutlich an zufälligen Stellen in das Genom der Wirtszelle eingebaut; damit liegt das HI-Virus als sogenanntes Provirus vor. Seine Gene lassen sich nun durch die menschliche RNA-Polymerase II ablesen, wobei die LTRs als starke Promotoren wirken.
- Anmerkung: Von der Bindung des Virus an die Wirtszelle bis zur Integration des Genoms vergehen etwa zehn Stunden. Nach Einbau der viralen DNA können Jahre vergehen, bis die Produktion der viralen Bestandteile effektiv beginnt und die Zelle schließlich unter Freisetzung neuer Viren lysiert wird. Es ist bis jetzt nicht verstanden, auf welchem Wege diese sogenannte Latenzperiode beendet wird, man schreibt jedoch dem Transkriptionsfaktor NFκB eine Funktion bei der Auslösung produktiver Zyklen zu.
Auch in der Latenzzeit kann das Immunsystem kompromittiert sein; die Ursache hierfür ist ebenfalls bisher unbekannt. Beim Befall größerer Mengen von immunkompetenten Zellen beginnt der Ausbruch von AIDS.
Biosynthese viraler Proteine
Nach Beendigung der Latenzphase beginnt die Wirtszelle mit der Transkription viraler Gene, wobei die vom HIV-Genom kodierten Regulatorproteine und wirtseigene Transkriptionsfaktoren an der Regulation beteiligt sind.
Von der gebildeten mRNA werden die viralen Proteine translatiert und posttranslational modifiziert: Zelleigene Glykosyltransferasen sorgen für die Glykosylierung der Hüllproteine gp41 und gp120, während die viruseigene Protease die beiden Proteine aus dem Vorläuferprotein gp160 freisetzt. Sie spaltet auch das Genprodukt von pol in die drei Enzyme Protease, Integrase und reverse Transkriptase
Assembling und Reifung
Aufgrund noch nicht verstandener Mechanismen bilden sich an bestimmten Stellen der Zellmembran Komplexe viraler Proteine, die die Ausknospung der Membran verursachen. Schließlich entstehen reife Viruspartikel, die alle für einen weiteren Infektionszyklus benötigten Proteine enthalten.
Durch Lyse der Wirtszelle werden sie wieder ins Blut freigesetzt und binden erneut an CD4+-Zellen.
Resistenz
In Patienten mit einer bestimmten Mutation des Zytokinrezeptors CCR5 kann sich das HI-Virus nur schwer replizieren, da die Virusfusion erschwert ist. Bei heterozygoten Merkmalsträgern ist die Infektion erschwert, bei homozygoten Personen scheint eine natürliche Resistenz gegen HIV vorzuliegen.
Immunsuppression
Die genauen Mechanismen der Wirkung von HIV auf das Immunsystem sind noch weitgehend unbekannt. Man weiß, dass nach einer Infektion mit dem HI-Virus das Immunsystem über die primäre Immunantwort in der Lage ist, die Viruslast im Blut nahezu auf null zu senken; allerdings überdauern HI-Viren in T-Helferzellen und anderen Zellen und entziehen sich auf nicht genau verstandene Weise dem Immunsystem: Vermutlich exprimieren die infizierten Zellen keine viralen Proteine auf ihrer Oberfläche und können damit nicht als befallen erkannt werden. Die sekundäre Immunantwort läuft aufgrund der T-Zell-Hemmung nur schwach ab und spielt daher bei der Abwehr von HIV nur eine untergeordnete Rolle.
Paradoxerweise sinkt die Anzahl der T-Helferzellen im Blut stärker ab, als man den Virustitern nach erwarten dürfte. Man vermutet, dass es neben dem Befall dieser Zellen durch das HI-Virus noch andere Mechanismen gibt, die zu einer Suppression der T-Zell-vermittelten Immunantwort führen.
Nachweis
Der labormedizinische Nachweis des HI-Virus kann mittels verschiedener Methoden erfolgen. So erfolgt i.d.R. zunächst ein HIV-Suchtests, bei dem es sich um einen ELISA-basierten, kombinierten Antikörper-/Antigennachweis aus dem Blut handelt. Ist dieser positiv, wird er nachfolgend durch einen Westernblot-basierten Bestätigungstest verfiziert.
Alternativ kann die virale RNA mittels RT-PCR nachgewiesen werden. Dieses Verfahren hat eine hohe Spezifität und Sensitivität, wird aber aus Kostengründen nicht routinemäßig angewandt.
Es sind ebenfalls freiverkäufliche Schnelltests verfügbar, die auch von Laien zu Hause durchgeführt werden können.
Detaillierte Informationen zum Erregernachweis finden sich im Artikel HIV-Infektion.
Meldepflicht
Gemäß § 7 des Infektionsschutzgesetzes (IfSG) besteht in Deutschland eine anonymisierte Meldepflicht für das diagnostizierende Labor. In der Schweiz sind vom Labor Vornamenscode, Wohnort, Geburtsdatum und Geschlecht des bzw. der Infizierten zu melden. In Österreich ist der Labornachweis einer HIV-Infektion nicht meldepflichtig.
Quiz
Bildquelle
- Bildquelle für Flexikon-Quiz: © Anna Shvets / Pexels