Desoxyribonukleinsäure








Trainier deine Lernmuskeln!
Mit Flash Cards, Quiz und mehr
LoslegenSynonyme: DNS, DNA, Thymonukleinsäure
Englisch: deoxyribonucleic acid, DNA
Definition
Die Desoxyribonukleinsäure, kurz DNA, ist ein Makromolekül, das in Form einer Doppelhelix aus 4 verschiedenen Desoxyribonukleotiden aufgebaut ist. Die DNA enthält die Gene, die den Aufbau aller Proteine kodieren und somit fast alle für die Zellfunktion notwendigen Informationen enthalten.
Nomenklatur
Im wissenschaftlichen Sprachgebrauch verwendet man statt dem deutschen Akronym DNS meist die englische Abkürzung DNA (deoxyribonucleic acid). Diese Abkürzung sollte auch im deutschen Schrifttum verwendet werden, um Verwechslungen mit dem Domain Name System (DNS) des Internets zu vermeiden.
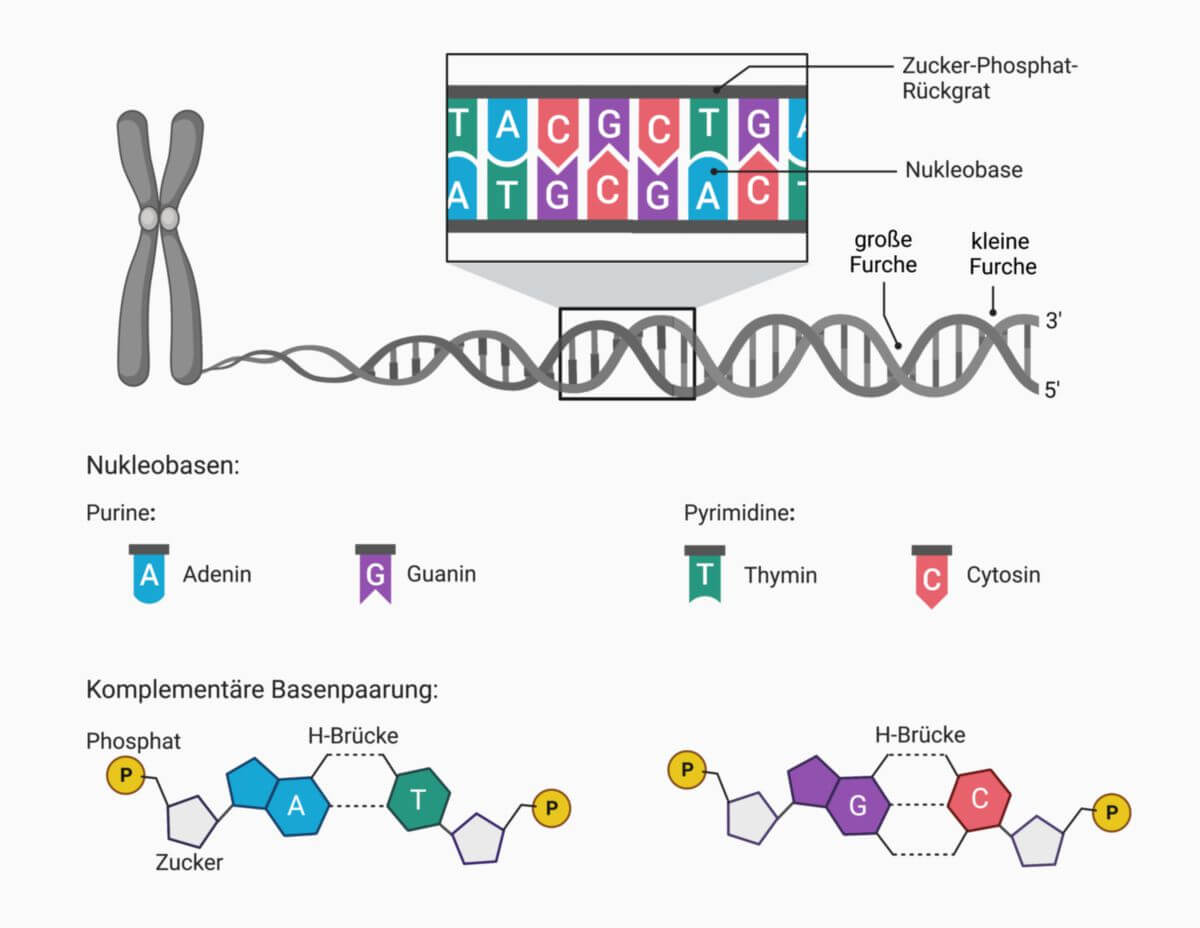
Aufbau
Die menschliche DNA ist aus zwei gegenläufigen DNA-Einzelsträngen aufgebaut. Jeder Einzelstrang hat ein 5'- und ein 3'-Ende. Am 5'-Ende sitzt ein Phosphatrest, am 3'-Ende eine OH-Gruppe.
Die Stränge formen eine Strickleiterstruktur, bei der die zwei Holme der Leiter um eine gedachte Achse schraubenförmig gewunden sind. Dadurch bildet sich die charakteristische Doppelhelix. Die beiden Holme der Strickleiter werden aus Hunderttausenden sich abwechselnder Zucker- (Desoxyribose) und Phosphatreste gebildet, die innerhalb jedes DNA-Einzelstrangs über kovalente Bindungen miteinander verknüpft sind. Dabei handelt es sich um Phosphodiesterbindungen, da die mit dem C3'-Atom eines Desoxyribosemoleküls veresterte Phosphatgruppe eine weitere Esterbindung mit der OH-Gruppe am C5'-Atom eines anderen Desoxyribosemoleküls eingeht. Durch diese Verknüpfung kommen die spezifischen 5'- bzw. 3'-Enden der DNA zustande.
Jede Sprosse der Strickleiter besteht aus zwei Nukleinbasen, die je über eine N-glykosidische Bindung mit einem Desoxyribosemolekül und Wasserstoffbrücken miteinander verbunden sind und so dafür sorgen, dass die beiden Holme auch im schraubenförmigen Zustand der Strickleiter verknüpft bleiben und im gleichen Abstand nebeneinander liegen. Insgesamt gibt es in der DNA vier verschiedene organische Basen: Adenin, Cytosin, Guanin und Thymin, die gewöhnlich mit den Anfangsbuchstaben A, C, G und T abgekürzt werden. Die Basenpaare werden von den jeweils komplementären Basen Adenin und Thymin sowie Cytosin und Guanin gebildet. Zwischen Adenin und Thymin bilden sich dabei zwei Wasserstoffbrücken aus. Cytosin und Guanin sind über drei Wasserstoffbrücken miteinander verknüpft. Zwei benachbarte Basen stehen in einem Winkel von 36° und mit einem Abstand von 0,34 nm zueinander.
Die DNA ist also aus einer Vielzahl von vier verschiedenen Nukleotiden "zusammengesteckt", die in einem DNA-Einzelstrang in beliebiger Reihenfolge aneinander gebunden werden können und sich dadurch unterscheiden, dass sie jeweils nur eine von vier möglichen organischen Basen enthalten.
Genetischer Code
Jeweils drei Basen, die in einem DNA-Einzelstrang direkt hintereinander liegen, bilden ein so genanntes Basentriplett oder Codon. Jedes Basentriplett steht für eine von 20 Aminosäuren, aus denen die Proteine aufgebaut sind. Die Reihenfolge der Basen - und damit der Basentripletts - bestimmt die Reihenfolge der Aminosäuren in den Proteinen. Dadurch wird der Aufbau der Proteine mit Hilfe der Basensequenz innerhalb der DNA beschrieben. Die Information der DNA wird bei der Proteinbiosynthese zunächst durch die Transkription in mRNA-Moleküle überschrieben. Die von der mRNA übermittelte Information wird dann durch Translation am Ribosom in eine Polypeptidkette übersetzt.
siehe Hauptartikel: genetischer Code
Konformationen
Die DNA-Doppelhelix kommt in drei unterschiedlichen Konformationen vor:
- B-DNA: Die B-Konformation ist die am häufigsten in vivo vorliegende Konformation. Die Helix dreht sich nach rechts, wobei pro Umdrehung ~ 10,5 Basenpaare (bp) verbaut sind. Das ergibt eine Hubhöhe von 3,4 nm. Die Wasserstoffbrückenbindungen stehen annähernd im rechten Winkel zur gedachten Helixachse.
- Z-DNA: In vivo macht diese Form nur einen sehr kleinen Anteil aus. Die Helix ist linksgängig mit 12 Basenpaaren pro Windung.
- A-DNA: Die A-Konformation tritt in Folge von Dehydrierung auf (z.B. bei getrockneter DNA). Die Doppelhelix weist pro Windung 11 Basenpaare auf und ist rechtsgängig.[1]
Kondensation
Bei einer diploiden menschlichen Zelle mit etwa 6,4x109 Basenpaaren, hätte die DNA bei linearer Anordnung eine Länge von rund 2 Metern. Um eine platzsparende Anordnung im Zellkern zu gewährleisten, muss das Molekül mit speziellen Hilfsproteinen, den Histonen, verpackt werden.[2] Im Zuge der Kondensation zu Chromatin wird die DNA dabei um Histonoktamere gewunden, es bilden sich die sogenannten Nukleosomen. Je nach Packungszustand kann sie als dicht gepacktes Heterochromatin oder lose gepacktes Euchromatin vorliegen, welche für Transkriptionsfaktoren zugänglich ist.
DNA-Replikation
Die DNA ist in der Lage, sich mit Hilfe von Enzymen selbst zu verdoppeln. Sie wird nach dem so genannten semi-konservativen Prinzip repliziert. Die doppelsträngige Helix wird zunächst durch das Enzym Helicase aufgetrennt. Ein Einzelstrang dient als Matrize für den zu synthetisierenden komplementären Gegenstrang, d.h. die replizierte DNA besteht jeweils aus einem alten und einem neu synthetisierten komplementären Einzelstrang. Der Vorgang der DNA-Synthese, d.h. die Bindung der zu verknüpfenden Nukleotide, wird durch Enzyme aus der Gruppe der DNA-Polymerasen vollzogen. Ein zu verknüpfendes Nukleotid muss in der Triphosphat-Verbindung - also als Desoxyribonukleosidtriphosphat - vorliegen. Durch Abspaltung zweier Phosphatteile wird die für den Bindungsvorgang benötigte Energie frei.
Im Bereich der durch das Enzym Helicase gebildeteten Replikationsgabel, d.h. zweier auseinander laufender DNA-Einzelstränge, markiert zunächst ein RNA-Primer den Startpunkt der DNA-Neusynthese. An das RNA-Molekül hängt die DNA-Polymerase dann ein zum Nukleotid des alten DNA-Einzelstrangs komplementäres Nukleotid, daran wieder ein weiteres neues passendes Nukleotid usw., bis die DNA wieder zu einem Doppelstrang komplettiert wurde. Dies geschieht an beiden geöffneten Einzelsträngen.
Die Verknüpfung der neuen Nukleotide zu einem komplementären DNA-Einzelstrang verläuft nur in 5'→3' Richtung, d.h. kontinuierlich den alten 3'→5'-Strang entlang in Richtung der sich immer weiter öffnenden Replikationsgabel ohne Pause in einem Schritt durch. Die Synthese des zweiten neuen Stranges am alten 5'→3'-Strang dagegen kann nicht kontinuierlich in Richtung der Replikationsgabel, sondern nur von dieser weg ebenfalls in 5'→3'-Richtung erfolgen. Die Replikationsgabel ist aber zu Beginn der Replikation nur ein wenig geöffnet, weshalb an diesem Strang - quasi in "unpassender" Gegenrichtung - immer nur ein kurzes Stück neuer komplementärer DNA entstehen kann.
Da hier jeweils eine DNA-Polymerase nur ca. 1.000 Nukleotide verknüpft, ist es notwendig, den gesamten komplementären Strang stückchenweise zu synthetisieren. Bei etwas weiter geöffnetem Zustand der Replikationsgabel lagert sich daher ein neuer RNA-Primer wieder direkt an der Gabelungsstelle an den DNA-Einzelstrang an, und die nächste DNA-Polymerase beginnt - sich von der Replikationsgabel entfernend - erneut ca. 1.000 Nukleotide an den RNA-Primer zu hängen. Dieser Vorgang wiederholt sich kontinuierlich, so dass der komplementäre DNA-Strang nach und nach etappenweise entsteht. Bei der Synthese des 3'→5'-Stranges wird also pro DNA-Syntheseeinheit jeweils ein neuer RNA-Primer benötigt. Primer und zugehörige Syntheseeinheit bezeichnet man als Okazaki-Fragment.
Die für den Replikationsstart benötigten RNA-Primer werden enzymatisch abgebaut. Dadurch entstehen Lücken im neuen DNA-Strang, die durch spezielle DNA-Polymerasen mit Nukleotiden aufgefüllt werden. Zum Abschluss verknüpft eine DNA-Ligase die noch nicht miteinander verbundenen neuen DNA-Abschnitte zu einem einzigen, langen, komplementären Einzelstrang.
Nach Abschluss der Replikation wurden also zwei DNA-Einzelstränge in etwas unterschiedlicher Weise jeweils wieder zu einem Doppelstrang ergänzt. Aus einem DNA-Molekül sind somit zwei entstanden.
DNA-Reparatur
Aufgrund ihres komplexen Aufbaus wird die DNA durch physikalische und chemische Einflüsse leicht beschädigt. Die Anzahl der DNA-Schäden liegt im Bereich von mehreren Zehntausend Einzelvorfällen pro Zelle pro Tag. Deshalb gibt es eine Vielzahl verschiedener Enzyme, die ständig mit der DNA-Reparatur beschäftigt sind. Stellt diese Reparatur den Ausgangszustand nicht wieder 1:1 her, entstehen Mutationen, die einen unterschiedlich starken Einfluss auf die weitere Funktion der Zelle haben.
Während einige Mutationen still bleiben, ziehen andere Funktionsstörungen im Zellstoffwechsel oder im Extremfall die Apoptose der Zelle nach sich. Mutationen der sogenannten Protoonkogene können zu einer malignen Transformation der Zelle führen.
Gelegentlich sind Mutationen der DNA aber auch von Vorteil. Sie bilden dann den Ausgangspunkt für die Veränderung von Lebewesen im Rahmen der Evolution.
Formen
DNA tritt in verschiedenen Formen auf, die meist durch Akronyme bezeichnet werden. Die wichtigsten sind:
- dsDNA: doppelsträngige DNA
- ssDNA: einzelsträngige DNA
- mtDNA: mitochondriale DNA
- Tumor-DNA: DNA von Tumorzellen
- cfDNA: zellfreie DNA in Körperflüssigkeiten
- cccDNA: ringförmig geschlossene DNA
Geschichte
Die Struktur der DNA wurde inkremental von mehreren Forschungsgruppen aufgedeckt. Beteilgt waren unter anderem AlecTodd, Rosalind Franklin, Raymond Gosling, Linus Pauling, Robert Corey, James Watson und Francis Crick. Watson und Crick erhielten 1962 den Nobelpreis für Physiologie oder Medizin.
Literatur
- James D. Watson: Die Doppelhelix. Rowohlt-Taschenbuch 1997
- James D. Watson: Gene, Girls und Gamov. Erinnerungen eines Genies. Piper 2003 ISBN 3-492-04428-X
- Ernst Peter Fischer: Das Genom. Fischer-Taschenbuch 2002
- Ernst Peter Fischer: Am Anfang war die Doppelhelix. James D. Watson und die neue Wissenschaft vom Leben. Ullstein 2003 ISBN 3-550-07566-9
- T.A. Brown: Moderne Genetik. 2. Aufl. Spektrum Akademischer Verlag, 1999
- James D. Watson, M. Gilman, J. Witkowski und M. Zoller: Rekombinierte DNA. 2. Aufl. Spektrum Akademischer Verlag 1993