Doppelstrangbruch






Trainier deine Lernmuskeln!
Mit Flash Cards, Quiz und mehr
LoslegenEnglisch: double-strand break, DSB
Definition
Als Doppelstrangbruch, kurz DSB, wird die Durchtrennung beider Nukleotidketten der DNA bezeichnet. Er gilt als die schwerwiegendste Form des DNA-Schadens.[1]
Ursachen
Doppelstrangbrüche können durch exogene und endogene Faktoren entstehen:
- Ionisierende Strahlung (z.B. Röntgen- oder Gammastrahlung)
- freie Radikale bzw. Reaktive Sauerstoffspezies
- Zusammenbruch der Replikationsgabel während der DNA-Replikation
- bestimmte Zytostatika (z.B. Topoisomerase-Inhibitoren, Bleomycin)
- mechanische Kräfte
- programmierte Doppelstrangbrüche (z.B. bei der V(D)J-Rekombination und der Meiose)
Biologische Bedeutung
Bei einem Doppelstrangbruch handelt es sich um den schwerwiegendsten DNA-Schaden. Kann die Zelle diesen nicht reparieren, ist die DNA-Replikation nicht mehr möglich und der Zelltod (z.B. durch Apoptose oder mitotische Katastrophe) wird eingeleitet. Ein Doppelstrangbruch trägt immer das Risiko einer genetischen Mutation, da nicht jeder Reparaturmechanismus fehlerfrei ist. Dadurch erhöht er auch das Entartungsrisiko der Zelle (Karzinogenese), beispielsweise wenn das Tumorsuppressorprotein p53 von einer inaktivierenden Mutation betroffen ist. Diese zentrale Rolle wird therapeutisch genutzt: Strahlentherapie und viele Zytostatika wirken über die Induktion irreparabler Doppelstrangbrüche, und Tumoren mit gestörter homologer Rekombination (z.B. BRCA1-/BRCA2-mutierte Karzinome) sind gegenüber PARP-Inhibitoren empfindlich.[2]
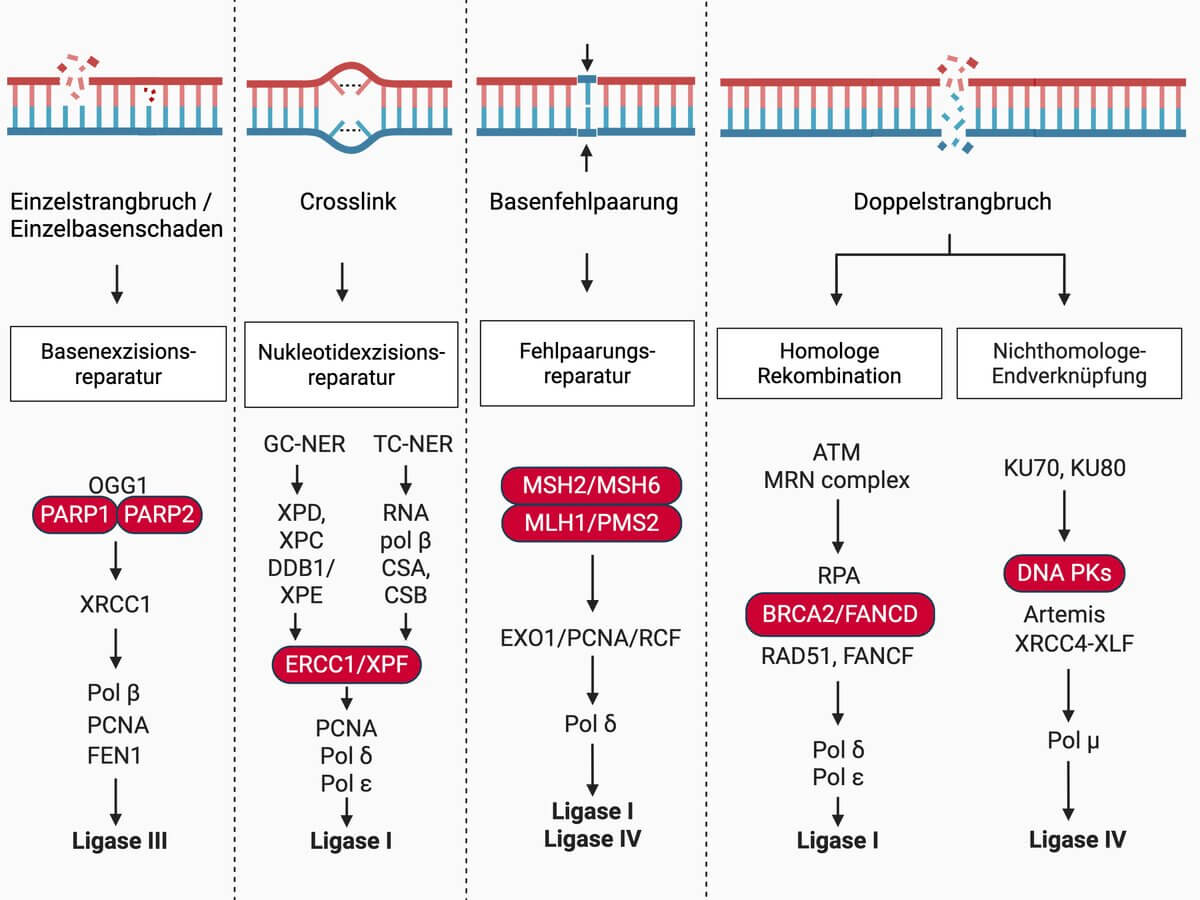
Reparatur
Die Wahl des Reparaturwegs hängt vom Zellzyklus und der Prozessierung der freien DNA-Enden ab.[3]
siehe auch: DNA-Reparatur
Homologe Rekombination
Die homologe Rekombination ist weitgehend fehlerfrei und in Säugetierzellen auf die späte S-Phase und die G2-Phase des Zellzyklus sowie auf die Meiose beschränkt, da sie ein Schwesterchromatid als Matrize benötigt. Entsprechend der Sequenz dient das Schwesterchromatid als Vorlage für die Auffüllung der Lücke. Beteiligt sind unter anderem die Proteine Rad51, BRCA2 und PALB2.[3]
Nicht-homologe Endverknüpfung
Die nicht-homologe Endverknüpfung (NHEJ) kommt bei Säugern und höher entwickelten Mehrzellern vor. Die DNA-Enden werden durch Reparaturenzyme nach Entfernung der geschädigten Nukleotide verknüpft. Beteiligt sind Ku-Proteine, welche die Enden der Doppelstrangbrüche erkennen und markieren. An die markierten Enden lagern sich die DNA-abhängige Proteinkinase (DNA-PK) und der Komplex XRCC4/DNA-Ligase IV an. Der Prozess läuft ohne homologe Matrize ab, dadurch wird nicht immer die richtige DNA-Sequenz hergestellt. In Säugetierzellen werden Doppelstrangbrüche vorrangig durch NHEJ entfernt.[3]
Microhomology-mediated end joining
Das Microhomology-mediated end joining (MMEJ) nutzt kurze homologe Sequenzen (Mikrohomologien) beiderseits der Bruchstelle, die nach Resektion aneinanderlagern. Da der dazwischenliegende Bereich verloren geht, ist der Mechanismus stark fehlerbehaftet und mit Deletionen verbunden.[3]
Single strand annealing
Das Single strand annealing (SSA) kommt bei Säugetieren vor. Die DNA-Enden werden bis zu flankierenden homologen Repeat-Sequenzen zurückgeschnitten, die anschließend aneinanderlagern. Der dazwischenliegende Abschnitt geht dabei verloren, sodass SSA immer mit einem Substanzverlust (Deletion) einhergeht.
Quellen
- ↑ Chatterjee N, Walker GC. Mechanisms of DNA damage, repair, and mutagenesis. Environ Mol Mutagen. 2017;58(5):235-263.
- ↑ Tan J et al. Double-strand DNA break repair: molecular mechanisms and therapeutic targets. MedComm (2020). 2023;4(5):e388.
- ↑ 3,0 3,1 3,2 3,3 Scully R, Panday A, Elango R, Willis NA. DNA double-strand break repair-pathway choice in somatic mammalian cells. Nat Rev Mol Cell Biol. 2019;20(11):698-714.